
|
Environmental risk factors
Environmental risk factorsEnvironmental risk factors and developmental windows of disease and disease prevention
At every stage of their life – from conception to death – organisms are exposed to a multitude of environmental factors, some of which are associated with severe health risks. Current research is now attempting to clarify the significance of particularly sensitive periods of the development of organisms, known as “developmental windows of disease”. Within these windows, there is an increased chance of specific types of changes to occur which interfere with genetically-determined development processes. The resulting reprogramming can increase the risk of the development of metabolic diseases, the impairment to reproductive health, or even the 
The development of a mammalian species from early foetal stages to adulthood is genetically determined. The precise course of this developmental programme is also subject to numerous environmental influences, with toxins, stress, behavioural factors and nutrition being the primary examples (Fig. 1). Nutrition exerts its influence on the organism via three pathways: while starvation and overeating (especially if this leads to excess weight) represent more general factors, bioactive compounds in food tend to have a highly specific effect on individual molecular structures, such as receptors or enzymes. The influence of environmental factors on the unfolding programmes of developmental biology is so potent that it may have a lifelong impact on the health of the individual. Modifications of the epigenom make some environmental influences inheritable: as a consequence, the health of subsequent generations can be influenced, a phenomenon termed the “developmental programming of disease”.

Fig. 1: Environmental factors and health. Environmental factors such as stress, toxins and changes in behaviour and nutrition are capable of influencing health later in life or the health of subsequent generations by modulating processes in developmental biology. The modulation of epigenetic mechanisms in particular thereby appears to play an important role. The influence of nutrition is twofold: we need to distinguish between the effects that are triggered by the availability and (calorific) quality of food sources (dearth/plenty) and effects produced by individual compounds contained in food products – which, like pharmaceuticals, affect highly-specific molecular target structures such as receptors or enzymes.
Environmental factors trigger influences over several generations From a mechanistic perspective, environmental factors appear to reprogram processes in developmental biology primarily by means of epigenetic mechanisms (for an overview, see Gerhäuser [1]). These programming events not only influence the generations directly affected but can also impact on their progeny, even though these individuals have had no direct exposure to the environmental factor – such as a chemical. For a pregnant individual, in the worst-case scenario the exposure to an environmental factor may directly affect the mother, the fetus and the priomordial germ cells of the fetus (i.e. the F0, F1 and F2 generations). If the resulting effect are not only detectable in the mother but in the F1 and/or F2 generation it is referred to as a multigenerational effect. If the effect persists into the F3 or a later generations – i.e. into a generation without any direct (e.g. chemical) exposure – it is termed a transgenerational effect. Both types of effect (multi- and transgenerational) have been indisputably demonstrated in animal studies for materials known as “endocrine disrupting chemicals” (EDCs), in relation to impairments affecting male reproductive health. The studies showed how the epigenome of the germline cells was reprogrammed, resulting in changes of processes within development and in epigenetic alterations that were passed on to male descendants via the male germline [2, 3]. These findings raise the question of the relevance of such mechanisms for human Development. Answers are to be found by looking at the effects from the Dutch famine in the winter of 1944/45. Researchers from Leiden University followed-up both the fate of mothers pregnant at the time and their children. The results tell us that the daughters of mothers affected by starvation have significantly elevated risks of suffering from a range of health problems, including metabolic disorders (such as obesity, impaired glucose tolerance and high blood pressure), neuronal disorders or hormone-dependent tumours such as breast cancer. These effects remain in force for their grandchildren. These examples demonstrate that environmental factors (here: caloric restriction) have the capacity to impact on processes of development thereby influencing the risk of developing a disease later in life and in subsequent generations. Since these results were published, epidemiological surveys and animal studies have provided some preliminary indications that bioactive compounds in foodstuffs may also have the capacity to prevent disorders by the modulation of the developmental programming [4, 5]. Relevant biological windows While we know that environmental factors can reprogram the trajectory taken by processes in developmental biology, the question of timing is also of crucial importance. The question is does the organism respond to the same degree of sensitivity in all stages of development from conception to adulthood – or are some stages especially sensitive and “fragile”? In analysing the consequences of the abovementioned famine in more detail, we can see that the observed effects behave similarly to the teratogenic effects from drugs such as Thalidomide, i.e. they correlate with specific stages in pregnancy rather than being general and are strictly dependent on when the mother suffered malnutrition. An elevated risk of obesity for the child correlated with a period of starvation of the mother in the first trimester of pregnancy, while a risk of impaired glucose tolerance was associated with hunger in the second trimester. The risk for high blood pressure arose when malnutrition occurred in the last trimester [4, 5]. In summary, we can say that: a) there are a number of “fragile” windows for the programming of adverse effects on health by environmental factors in the early stages of development; and that b) these windows of fragility appear to be specific to organs or processes. The largest risk factor for an increased risk to develop a malignant tumour of the female mammary gland appears to be the quantity of autologous oestrogens and substances that mimic oestrogens over the course of the womans lifetime. Exposure to these hormones does not seem to pose an equal level of risk for a woman at every stage of her life. In humans, particularly critical phases for oestrogen exposure discussed in the literature to date include early foetal development, development of the mammary glands at puberty and the involution of the mammary gland following cessation of breastfeeding or involution of the breast during climacteric transition (Fig. 2 and [6]). Animal studies have also revealed an additional critical phase for exposure, namely the neonatal development of the mammary gland.
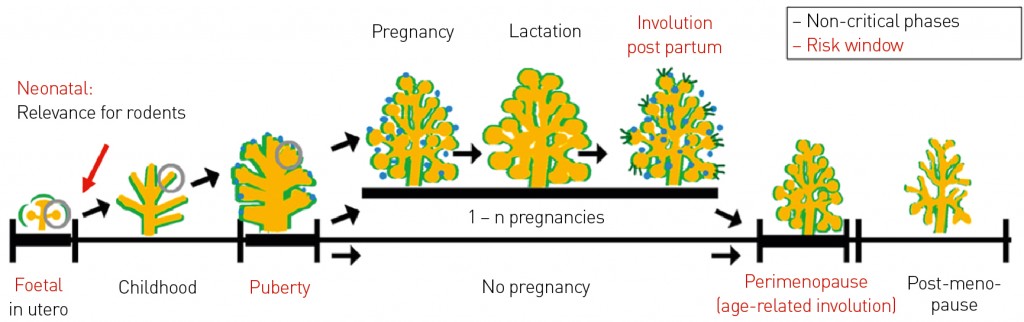
Fig. 2: Hypothesis of developmental window for breast cancer risk. Schematic diagram of the development and lifecycle for the female mammary gland. The periods of pregnancy, lactation and post-lactational involution distinguish mothers from women who have never been pregnant. The four windows with particular relevance for the risk of breast cancer are marked in red, and include parts of foetal and pubertal development, as well as involution processes triggered by the cessation of breastfeeding or decreasing hormone production in the ovaries. Modified after [6].
Xenoestrogens from the environment Due to these influences, “environmental oestrogens” have been a particular focus of research. These xenobiotics with oestrogen-mimicking properties include both industrial chemicals and substances of plant or fungal origin (phyto/myco-oestrogens). Since plant sources are particularly rich in phyto-oestrogens, extracts are prepared from these sources and offered commercially as alternative for the treatment of climacteric/menopausal complaints. Such substances include isoflavones from soybean, red clover and kudzu, as well as hop-derived naringenins. Our lab is currently investigating these kinds of extracts – and single chemicals derived thereof – in terms of their efficacy (e.g. prevention of osteoporosis, suppression of vasomotor disorders) and safety (exclusion of any risk for breast cancer genesis or endometrial hyperplasia) in menopausal applications thereby using preclinical experimental models. As detailed above, we have been working for many years now on the hormonal effectiveness of ingredients from foodstuffs and preparations of medicinal plants – including isoflavones from legumes. Yet what is the rationale to switch from investigations of the molecular and cellular effects of soybean preparations and their bioactive principles to a topic of developmental biology namely the dietary modification of high-risk windows for mammary gland development? The idea to deploy soybean preparations for the treatment of hormone-based symptoms – such as menopausal complaints – originated in observations made by H. Adlercreutz [7]. Adlercreutz’s research compared groups of European and Asian subjects, showing that the level of isoflavones (the active ingredient in soybean) in urine was inversely correlated to the risk of breast cancer in these groups. The causal relationship between the quantity of soy products consumed and the lowering of the risk of breast cancer development was substantiated only later by epidemiological data from the period 2009 to 2014 [8]. According to these findings, a lifelong dietary consumption of soy isoflavones appears to be associated with a lower risk for the development of breast cancer. While clinical trials investigating the use of preparations enriched with soy isoflavones by menopausal women have up to date provided no indications of side effects – such as changes to the density of the breast tissue, a number of animal studies have produced findings that indicate soy isoflavones are nonetheless capable of promoting the growth of human breast cancer cells, which were implanted into immunodeficient nude mice [9]. If we translate the above insights into the pharmacological perspective, this means that a lifelong dietary consumption of soy isoflavones is equivalent to a chronic low-dose exposure that may exert a beneficial effect towards breast cancer risk in adulthood. The consumption of dietary supplements enriched in isoflavones at the point in time of the menopausal transition, on the other hand, would correspond to a comparatively high-dose treatment in later life. The research questions that our team has been addressing in recent years have were delineated from both the observations described above, and the hypothesis that the development of tumours of the breast is also affected by exposure during particularly sensitive and therefore critical windows of time. These questions are: a) how does a lifelong soybean-rich diet affect the potency of the female sexual hormone oestradiol as a driving force in the carcinogenesis of hormone-dependent organs in the experimental model of the rat; and b) how does a potential modulation of oestradiol effects affect the development of tumours in an oestradiol-driven, experimental breast cancer model? Isoflavones modulate the activity of oestradiol Since the activity of oestradiol has a decisive role in the carcinogenesis of hormone-dependent organs, we performed an initial multi-generational feeding study with animal feed containing predefined quantities of soybean-derived isoflavones. Control animals received a control feed containing no phyto-oestrogens. Feeding with these diets began in the parent generation prior to mating and was sustained in the first generation of progeny throughout all critical windows of development (foetal, neonatal and pubertal phases). From the 97th day of life after birth, we used a procedure known as a “uterotrophic assay” to assess the extent to which the isoflavones in feed affected oestradiol responsiveness in the target organs of uterus and mammary gland. Key findings here were as follows: the sensitivity of the tissue of the uterus to oestradiol treatment was increased significantly [10], while the oestradiol-dependent stimulation of proliferation of the mammary gland epithelium was clearly attenuated by isoflavones [11]. The latter finding is clearly relevant when assessing the impact of soy isoflavones on the risk of breast cancer. Since values for bone mass and density were also elevated in the animals that received feed enriched with isoflavones, we may summarise the findings from these investigations as follows: a) continuous exposure to soy isoflavones reprograms the responsiveness of hormone-dependent organs to the female sexual hormone oestradiol; and b) the extent and the direction of the effect triggered by environmental factors on developmental programs is organ-selective – as is shown by the opposite nature of the effects manifested in the two primary target organs for oestradiol, namely uterus and mammary gland. Do isoflavones also modulate carcinogenesis of the mammary gland? In the human mammary gland, carcinogenesis is an oestradiol-dependent process in about 70?% of cases. This holds for both the initiation of carcinogenesis by genotoxic oestradiol metabolites and for the promotion of carcinogenesis of the mammary gland by oestradiol in a receptor-mediated mechanism. For this reason, our finding that isoflavones modulate the sensitivity of breast tissue to oestradiol is of particular relevance. The goal of our current work is to investigate whether and how isoflavones are also able to influence oestrogen-dependent carcinogenesis of the mammary gland. To answer this question, we require a suitable model in which development of breast cancer is dependent on oestradiol. Female ACI rats are especially suitable for this purpose. Artificial elevation of endogenous oestradiol levels in these animals greatly increases the incidence of breast tumours [12]. This protracted tumour experiment was carried out jointly by Dr Frank Möller and Dr Oliver Zierau in our research team. In technical terms, adapted the experiment on our feeding study described above with isoflavone-rich feed compared to isoflavone-free placebo feed. At the onset of puberty, an oestradiol releasing device was implanted and the experiment was conducted until postnatal day 285.Whole-mount preparations of the mammary gland (Fig. 3) were used to allow the authors to follow the progression of the process.
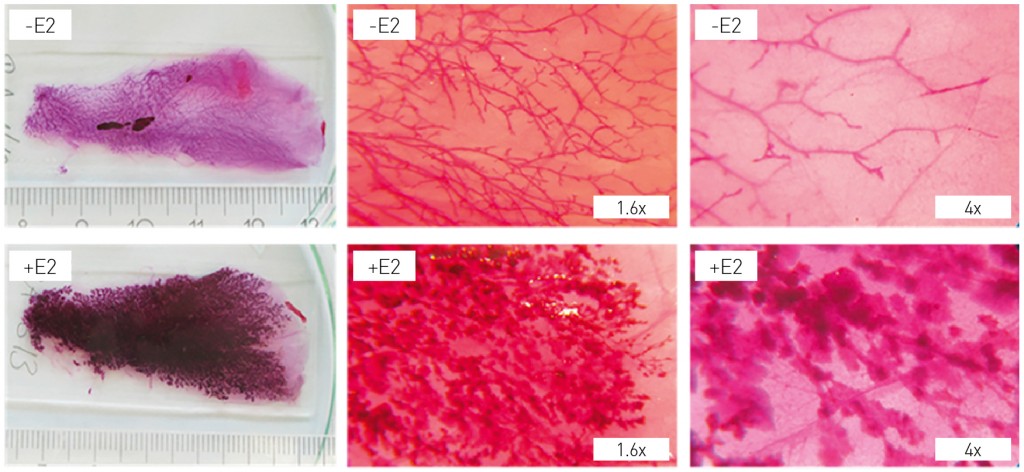
Fig. 3: Whole-mount preparations as indicators for changes in the mammary gland. Investigations using experimental tumour models are time-consuming. In many cases, potential changes in the breast due to experimental conditions can be observed at a relatively early stage in whole-mount preparations of the organ. The figure contrasts gland ducts in preparations of the breast with and without oestradiol stimulation (+E2/-E2). The figures were supplied by Dr Frank Möller (Molecular Cell Physiology and Endocrinology, Dept. of Biology, TU Dresden, Germany).
Results from this promising experiment have yet to be completed and therefore final, detailed data are not available yet. We can already report one extremely important and unambiguous finding, however: exposure to soy isoflavones throughout all of the sensitive phases of mammary gland development does modulate oestradiol-dependent carcinogenesis!
Bibliography Picture: © istockphoto.com| vm |
L&M int. 3 / 2015
Free download here: download here The Author:Read more articles online

|
Search:





