
|
From fat droplets in plant cells to novel foods
From fat droplets in plant cells to novel foodsFunctional nanoparticlesOccurring naturally in oilseeds, oleosomes are particles with special properties. Depending on the plant variety, their size ranges from microns down into the nanoscale. These particles, with their protein-functionalized surfaces, are structurally very stable indeed. This makes them relevant for fundamental research and pharmacology, and also for innovative applications in food science. Plant-derived fats: oilseeds and oil plants 
What do maize, soy, sesame, palm, sunflower, olive, peanut and pumpkin have in common? They are all termed “oil fruits” in agricultural jargon and – depending on the formulation of their fatty acids – have satisfied a huge range of culinary requirements as a source of vegetable oil for many hundreds of years. The botanist makes a further distinction, distinguishing the fruit or fruit pulp (olive, palm) from the seed or seeds, depending on the plant tissue from which the oil is actually isolated. Outside the realm of botany, few details were known until recently about how, why and where plants store their oil. A large body of research on these oil reservoirs can be found for oil seeds such as maize, sunflower seeds and soybeans. These reservoirs are known as “oil bodies” in the relevant literature or oleosomes (Fig. 1). A better understanding of these oleosomes can not only explain the Herculean efforts required by oil producers to extract the oil from the oil seed but also deliver a number of fundamental insights into the function and interaction of phospholipids with proteins. The typical size of an oleosome is at the nanoscale (diameter in soybeans approx. 300nm), and is essential for seed germination and growth. At the time of germination, when the plant is still unable to resort to photosynthesis, the stored neutral fats are broken down with enzymes into free fatty acids and ultimately monosaccharides, which are required for the construction of carbohydrates [1]. Since a smaller volume leads to an enlarged total surface area, the oleosomes offer a physically larger target to the fat-digesting enzymes (lipases). This is important, as it ensures the greatest amount of energy is available for the seedling during the short period of germination. To biosynthesize fats, plants make good use of their huge diversity of organelles (cell organs) and their enzyme apparatus. The biosynthesis of fatty acids takes place solely in the plastids, the most familiar of which are the green chloroplasts. According to endosymbiotic theory, these developed from cyanobacteria. Accordingly, their fatty acid constructor enzymes are of prokaryotic origin (descended from bacteria). This is the reason why plants can manufacture fatty acids that are essential for humans – i.e. which the human organism cannot manufacture itself. To do so, the plant uses enzymes such as -1,2-desaturase. This is a desaturating enzyme: it builds carbon/carbon double bonds into fatty acids: saturated fatty acids are turned into unsaturated ones – such as the essentially fatty acids linoleic acid (di-unsaturated) and linolenic acid (tri-unsaturated). Unsaturated fatty acids are not produced by plants merely for the convenience of the consumer or nutritional scientist, but play a key role in establishing the plant’s tolerance to cold weather. Thanks to their low melting point, these fatty acids remain liquid even at low temperatures. Cold-tolerant plants store greater amounts of phospholipids with unsaturated fatty acids in their cell membrane, thus increasing its fluidity (and lowering their crystallization temperature). Quite unlike a tropical plant such as the coconut: in the coconut, the tendency of unsaturated fatty acids to oxidize means saturated fatty acids tend to be stored, and the melting point is controlled merely by chain length. Accordingly, coconut oil is a highly saturated fat, but with short fatty acids. Thanks to insights obtained from plant physiology and cell biology, we now know how plants manage to distribute – i.e. emulsify – oil within a plant cell in the form of tiny lipid droplets (the oleosomes). In the plant cell, a nanoscale emulsifying process is effectively in place that has been fine-tuned by evolution over millions of years. At a specialized cell organelle (the endoplasmic reticulum), the ready-made fats manufactured by the enzymes accumulate together within this cell’s membrane (i.e. between the phospholipid lipid bilayer). This causes the membrane to balloon out into small droplets, which can then detach from the membrane [2]. The size of the droplet is predetermined by the oleosin proteins already mentioned, which simultaneously become stored in the newly-formed oleosome.
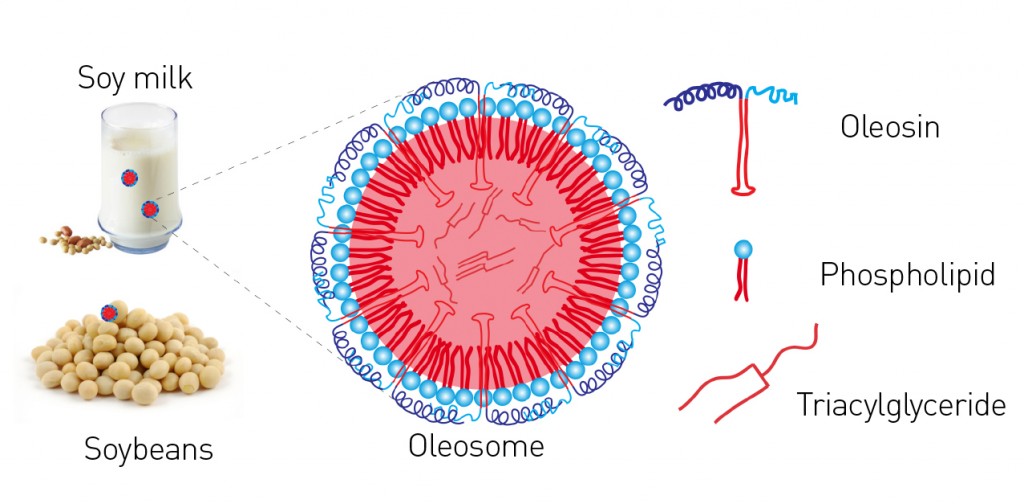
Fig. 1: Simplified illustration of an oleosome from oil seed, e.g. from soybean. Phospholipids form a stable layer but, unlike fat particles of animal origin (LDL and HDL), they do so without cholesterol. Additional stability is achieved via oleosins. These specially-formed proteins with a strongly lipophilic portion, which is bent into the shape of a "hairpin" (top right), are also deposited and act to stabilize the oil bodies. The hydrophilic portions of the protein consist of helices and unstructured polar regions.
Oleosins: special-purpose proteins These proteins, which stabilize the individual oleosomes, are unique in both their form and function. As an extreme simplification, an oleosin is like an umbrella: the upper part of the umbrella extends into water (and so is hydrophilic) and the handle is rooted in oil (hydrophobic) (see Fig. 1). Oleosins are very stable precisely because of this strong oil “anchor”: it consists of a sequence of about 70 hydrophobic amino acids, which form themselves into a “hairpin”-like structure. This sequence is the longest hydrophobic amino acid sequence known to occur naturally [2]. Another characteristic feature of oleosins is the “proline knot”, which contains three prolines (an amino acid known as a “helix breaker”) and forms the knot-like 180° turn of the hairpin. This medial hydrophobic sequence in the oleosins is identical in all plant seeds. Differences are in fact seen only in the edge sequences –the oleosins’ hydrophilic umbrellas – known as the N-terminal and C-terminal domains. These can consist of various hydrophilic domains such as an amphipathic a-helix, for example. The charge present in the outer umbrella portion of the oleosins and the repulsion of two individual oleosomes that results is not only primarily responsible for the stability of soy milk, but also the reason why seeds can retain their power to germinate for centuries at a time. This also explains why soy milk, tofu and soy whip are extremely efficient emulsifiers. Soy milk contains some of the strongest emulsifiers in existence: phospholipids, oleosins – i.e. proteins that contribute greatly to stability – and oil bodies, which all have their part to play at various orders of magnitude. This means the surface of the oleosomes is naturally “functionalized”: depending on the pH value of its surroundings, the surface bears either a positive or negative charge (Fig. 2). The net charge on the surface is positive below the isoelectric point and negative when above it. Accordingly, oleosomes have a slight positive charge in the acidic range of pH values around 4 that is of culinary relevance. They form stable emulsions and can be correspondingly encapsulated, e.g. with negatively-charged hydrocolloids.
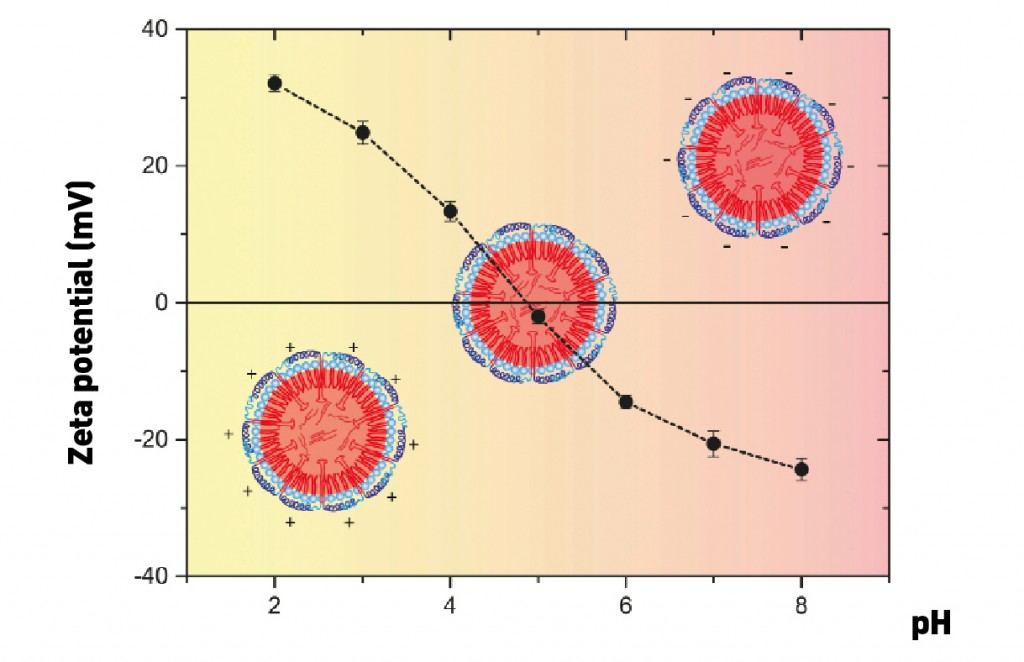
Fig. 2: Measurement of the zeta potential provides information about the surface charge of the oleosomes at various pH values [3].
Extraction Oleosomes are extracted from raw filtered soy milk in a multi-stage process using a centrifuge. This accelerates the formation of an oleosome “cream”, which is then separated from the soy milk’s remaining components. The pH value of the raw soy milk and of the medium used in the separate washing stages has a major influence on the degree of success in separating out the storage proteins. With a strongly basic pH value of 11, the separation of oleosomes from the storage proteins glycinin, ß-conglycinin and other allergens such as the thiol protease “Gly m Bd 30 K” is achieved [4]. Inside an oleosome: neutron scattering Although various models and simulations exist for the conformation of oleosins within the oleosome, their folding at the molecular level has not been definitively elucidated. It is also a difficult matter to demonstrate small changes in the conformation of the proteins (such as we expect at the beginning of thermal denaturing, for example) by macroscopic measurements of intact oleosomes. Neutron scattering offers us a promising approach to acquiring more precise insights in this context. Neutrons interact with the nuclei of atoms and have the special quality of exhibiting scattering differences when interacting with isotopes of the same element. This is especially pronounced in the case of hydrogen (H) and deuterium (D). This offers the possibility of suppressing or highlighting specific substructures within a sample by substituting hydrogen for deuterium to vary their scattering length densities. In the case of oleosomes, this gives us the option of dispersing them in mixtures with a range of various H2O/D2O ratios instead of normal water: this masks the normally dominant signal from the oil while boosting the weak but particularly interesting signal from the protein coat. This is illustrated by Figure 3, where the scatter signals from soy oleosomes are presented with the signals from a model emulsion – also based on soy and lecithin but protein-free (Intralipid) – in both pure H2O (0% D2O) and in 12%D2O. It can be seen that the signals of the two samples show virtually no differences for pure water, but differ significantly in the case of the 12% D2O mixture. This is because 12% D2O is, from a neutron’s perspective, virtually indistinguishable from soy oil, and the resulting signal is therefore dominated by the protein coat. In water, however, the oil signal is dominant. If we then compare the scattering curves with those calculated for certain structures and H2O/D2O ratios, we can make a statement about the structure of the sample. In the example shown, it can be readily seen that, while a simple sphere model can be used as an approximate description of both oleosomes and Intralipid in the case of pure water, this remains true only for the protein-free Intralipid in the case of the 12% D2O mixture. To be able to describe the signal generated by the oleosomes, it is necessary to use the more complex model of a sphere with a shell. By adjusting the model to fit the scattering curve, parameters such as oleosome size and shell thickness can be determined. This results in a diameter of 290nm and a shell thickness of 7nm, which is compatible with the above-mentioned model representation of the oleosome structure.

Fig. 3: Small Angle Neutron Scattering (SANS) applied to oleosomes and model emulsions clearly shows the oleosome's protein coat with measurement in 12% D2O.
Behavior at interfaces The basic behavior of oleosomes at interfaces is illustrated in Figure 4. If an oleosome reaches the air/water interface, the oleosins on the surface change their conformation as soon as they come into contact with air (which can be viewed as being hydrophobic). This causes the oil body to lose its stability: it ruptures and its components reconfigure themselves into new structures. In the process, the interface-active phospholipids arrange themselves at the surface as shown and the oil itself also collects at the surface. The behavior of the oleosins remains to be fully elucidated. Although the oleosins’ amphiphilic character also presents them with the opportunity of arranging themselves along the interface, micelle-like aggregates can also form (as shown), with which the hydrophobic partial sequences are enclosed by the hydrophilic sequences and can thus become water-soluble. An explanation will require further investigation (light scattering, spectroscopy, ...). This also makes oleosomes interesting in the field of cosmetics. In thickened aqueous solutions, they burst due to the slow evaporation of the solvent and thus release their oil reservoirs equally slowly.
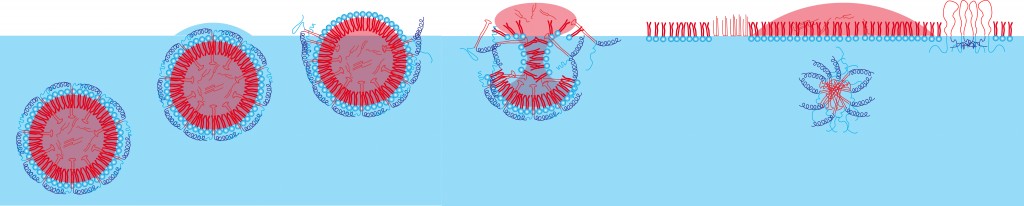
Fig. 4: Schematic diagram of the oleosome interface behavior, as illustrated by Langmuir trough experiments, combined with Brewster angle microscopy [4]. The oleosomes diffuse onto the surface. Once the hydrate water evaporates, the hydrophilic parts of the oleosins denature, the hydrophilic phospholipid heads reorient themselves, the oleosomes burst (hydrostatic pressure). At the air/water interface, various phases of fats, phospholipids and oleosins then form.
Oleosomes and innovative foods The functionalized surfaces of the nano- and microparticles can be used to bind the oil bodies into structured aggregates or to process them into solid and semi-solid components for subsequent use in novel foods. Some initial studies have already explored a number of possibilities. On the one hand, in the basic region and near the isoelectric point, for example, negatively-charged amino acids are predominant in the hydrophilic regions of the oleosins: these could be linked by Ca2+ ions, thus leading to a moderate improvement in stability (see Fig. 5). On the other hand, the hydrophilic parts of the protein could be permanently cross-linked with the enzyme transglutaminase at the surfaces. Transglutaminase requires calcium to develop its maximum level of activity: accordingly, maximal stabilization of the emulsion will result only from the interplay of the two substances.
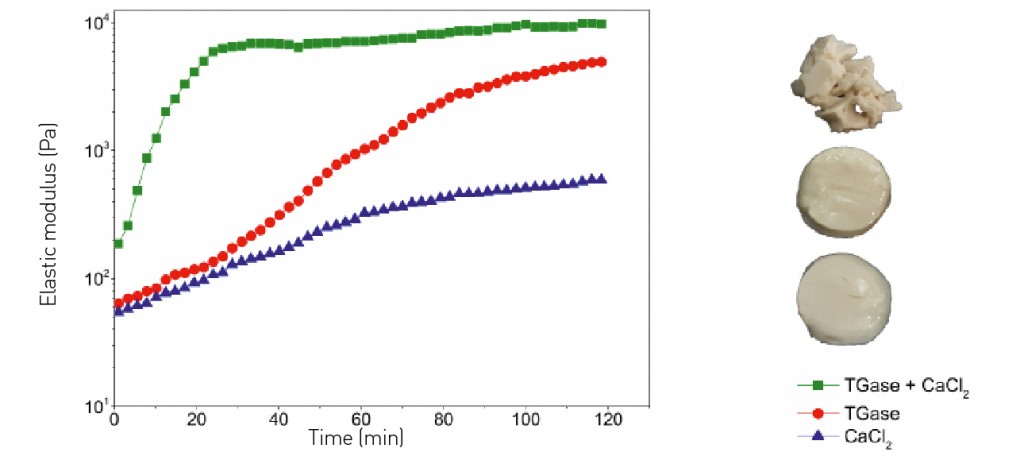
Fig. 5: Increased stability by cross-linking hazelnut oleosomes with calcium chloride, transglutaminase and a combination of the two. The texture is shown on the right-hand edge of the figure. The calcium gels are very soft, almost melting; the oleosomes permanently cross-linked with transglutaminase exhibit a considerably higher shear modulus. The texture is reminiscent of jelly. The combination of TGase and calcium (cofactor) leads to rapid cross-linking and the formation of fragile, yet soft, gels.
Oleosomes could therefore be used to develop protein-rich foods with an energy content that would be controllable via their fat percentage. This would also be of interest in geriatrics for the prevention and treatment of malnutrition – especially since the texture (and the ease with which food could be swallowed in the event of simultaneous dysphagia) could be precisely modified [5]. If we also up the ante by pointing out that both the surfaces and the oil reservoir can be loaded with bioactive substances of varying solubility (water- and fat-soluble vitamins, phenols, etc.), then oleosomes have every chance of becoming natural biofunctional carrier particles.
Bibliography Picture: istockphoto.com | Bogdanhoda |
L&M int. 4 / 2015
Free download here: download here The Authors:Read more articles online



|
Search:









